
| Entstehung: |
|
Die Haupt-Klone ("major clones"), die anhand ihrer unterschiedlichen Farbmuster bestimmbar sind
unterscheiden sich deutlich voneinander in mtDNA, Ploiditätswert und Allozymen. Die jeweiligen Haupt-Klone unterteilen sich nochmals in vom äußeren Erscheinungsbild her nicht erkennbar verborgene Unter-Klone("minor clones"), die sich durch leichte Abweichungen der Allozyme voneinander unterscheiden und warscheinlich durch Mutation innerhalb des jeweiligen Haupt-Klons entstanden sind (Bolger / Case, 1994). Die diploiden Klon-Linien sind aus Hybridisierung zwischen bisexuellen Spezies hervorgegangen. Bei den triploiden Klon-Linien geht man davon aus, dass sie durch Rückkreuzungen zwischen diploiden Klonen und Männchen der bisexuellen Elternspezies entstanden sind (Cole et al., 1988; Darevsky, 1992; Moritz, 1983; Radtkey et al., 1995; Volobouev et al., 1993; Quellen: Ota et al., 1989; Ota et al., 1996) Lange Zeit war unklar welche die bisexuellen Vorfahren sind, die bei den Hybridisierungen beteiligt waren, aus denen die verschiedenen Klone hervogegangen sind. 1995 ergaben Untersuchungen von Radtkey et al., dass Lepidodactylus lugubris aus einer Kreuzung zwischen Lepidodactylus moestus (beschrieben von Ota et al., 1995) und einer anderen unbeschriebenen Spezies (Lepidodactylus sp.) auf dem Arno Atoll (Marshall Inseln) entstanden ist, dass solche Kreuzungen mehrfach erfolgt sind und dass noch immer neue Klone entstehen. 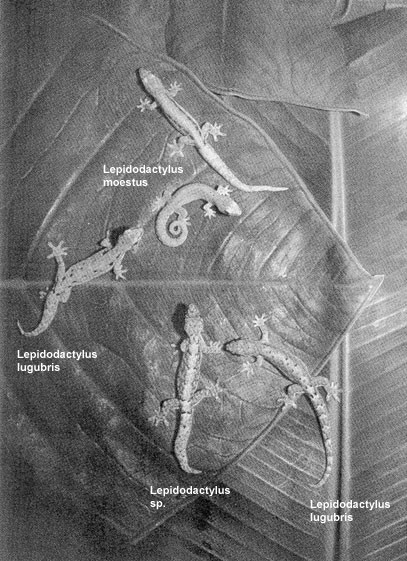
Zwei parthenogenetische Lepidodactylus lugubris Klone mit den beiden Eltern-Spezies. Zwei Farbwechsel von Lepidodactylus moestus, dem Vorfahren mütterlicherseits oben Mitte, der unbeschriebene Vorfahre väterlicherseits unten Mitte und zwei Lepidodactylus lugubris Klone Mitte links und unten rechts. Der Rückenmusterungs-Polymorphismus der mütterlichen Species wurde auf die Lepidodactylus lugubris Klone vererbt. Fotograf: J. Newsome. Quelle: Abbildung 1, Seite 147 aus Radtkey, R.R., S.C. Donellan, R.N. Fisher, C. Moritz, K.A. Hanley & T.J. Case, 1995. When species collide: the origin and spread of an asexual species of gecko. Proc. R. Soc. Lond. B259: 145-152 Boissinot et al. (1997) vermuteten ebenfalls, dass die Eltern-Populationen Lepidodactylus moestus und Lepidodactylus sp. vom Arno Atoll sind. Ihre Untersuchungen (1997) in Französisch Polynesien (Moorea Insel, Maiao Insel, Society Archipel, Rangiroa Atoll, Takapoto Atoll, Tuamoto Archipel) deuteten darauf hin, dass die bisexuelle Spezies auf Takapoto keine Eltern-Population der dortigen Klone darstellt aber möglicherweise zur Elternspezies gehört. Ein weiteres Ergebnis der Untersuchungen von Boissinot et al. (1997) belegt, dass Klon A unabhängig von Klon B, C und D entstanden ist und dass durch Hybridisierung zwischen dem diploiden Klon D und Männchen verschiedener anderer bisexueller Populationen die Klone B und C entstanden sein könnten. Moritz et al. (1993) geben unabhängiges Entstehen für die diploiden Klone 2nA, 2nB-I und 2nB-II an. Lt. Ihren Untersuchungen sind die triploiden Klone 3nB warscheinlich aus Kreuzung der diploiden Klone 2nB mit Männchen der bisexuellen Spezies entstanden und bei einem ähnlichen Ereignis mit Weibchen der diploiden Klone 2nA die triploiden 3nA Klone. Die Vielzahl der Linien von Lepidodactylus lugubris ist bis heute weltweit noch kaum erforscht. Hier die Beschreibungen einiger Linien (Ineich, 1987/1988; Ineich / Ota 1992; Yamashiro / Toda / Ota, 2000): |
| Einige Klone: | >> Seitenanfang |
|
Die Linien werden bestimmt mit Bezug auf die Beschreibungen von Ineich (1988). Diploider Klon A (Takapoto Atoll): Zwei Längsreihen bestehend aus 6 bis 8 (meist 7) dunklen Flecken vom Nacken bis zum Schwanzansatz, dorsale Grundfärbung hellgrau oder elfenbeinfarbig (Ineich, 1988; Ineich / Ota 1992) Triploider Klon B (Takapoto Atoll): Zahlreiche dunkle Flecken, besonders groß und insbesondere auf Nacken und Schwanzansatz paarig Dorsale Grundfärbung hellgrau oder elfenbeinfarbig. (Ineich, 1988; Ineich / Ota 1992) Triploider Klon C (Takapoto Atoll): Zwei Längsreihen von 6 bis 8 dunklen Flecken fom Nacken bis zum Schwanzansatz, zwei längliche dünne Striche im Nacken, dorsale Grundfärbung häufig rotbraun. (Ineich, 1988; Ineich / Ota 1992) Klon E, Ploidität unbekannt (Takapoto Atoll): Keine einheitlichen dunklen Muster, dorsale Grundfärbung rot-braun (Ineich, 1988; Ineich / Ota 1992) Diploider Klon D (Moorea Insel): Eine Reihe aus weniger als 7 einfachen Flecken nur im vorderen Bereich entlang der Rückenmitte (Ineich, 1988) |
|
Diploider Klon A (Haha-jima, Ogasawara Islands): Zwei Längsreihen bestehend aus sieben oder acht kleinen v-förmigen Mustern nur entlang der Mittellinie des Rückens (Yamashiro / Toda / Ota, 2000) Triploider Klon B (Kitadaitojima, Daito Inseln): Laterale schwarze Balken auf Nacken und Schwanzansatz, w-förmiges Zeichen auf dem Kopf, zwei dorsolaterale Reihen großer Halbmond-förmiger schwarzer Flecken (Yamashiro / Toda / Ota 2000) Triploider Klon C (Zamami-jima / Okinawa Inseln, Ryukyu Archipel): Dorsolaterale schwarze Balken auf Nacken und Schwanzansatz und schwach ausgeprägte w-förmige Zeichnung entlang der Mittellinie (Yamashiro / Toda / Ota 2000) Diploider Klon Da (Minamidaito-jima, Daito Inseln): Dorsales Muster bestehend aus lateralen schwarzen Balken auf Nacken und Schwanzansatz, Winkel und kurze ineinander übergehende Balken entlang der Mittellinie, ein relativ schwach ausgeprägtes w-förmiges Zeichen auf dem Kopf (Yamashiro / Toda / Ota, 2000). |
 Diploider Klon A (Haha-jima, Ogasawara Inseln) |
 Triploider Klon C (Zamami-jima / Okinawa Inseln, Ryuku Archipel) |
Bildtafel aus: 'Clonal composition of the Parthenoge- netic Gecko, Lepidodactylus lugubris, at the Northernmost Extremity of Its Range' von Saiko Yamashiro, Mamoru Toda und Hidetoshi Ota, 2000 |
 Diploider Klon Da (Minamidaito-jima, Daito Inseln) |
 Triploider Klon B (Kitadaitojima, Daito Inseln) |
 Triploider Klon B' (Minamidaito-jima, Daito Inseln) |
 Triploider Klon BI-1 (Minamidaito-jima, Daito Inseln) |
 Triploider Klon BI-2 (Minamidaito-jima, Daito Inseln) |
 Triploider Klon BI-3 (Minamidaito-jima, Daito Inseln) |
 Triploider Klon BI-4 (Minamidaito-jima, Daito Inseln) |
 Triploider Klon BI-5 (Minamidaito-jima, Daito Inseln) |
 Triploider Klon BI-6 (Minamidaito-jima, Daito Inseln) |
 Triploider Klon BI-7 (Minamidaito-jima, Daito Inseln) |
 Triploider Klon BI-8 (Kitadaito-jima, Daito Inseln) |
 Triploider Klon N (Minamidaito-jima, Daito Inseln) |
|
Diploide Gruppe Klone 2nA (Fiji/Hawaii/West-Pazifik): Bis auf Klon 2nA-III von Hawaii Musterung wie Klon A (Ineich, 1988), kleine gleichmäßige gepaarte Zeichen entlang der Rückenmitte. Klon 2nA-III auf Hawaii hat zusätzlich noch Zeichen seitlich des Nackens (Moritz et al., 1993). Diploide Gruppe Klone 2nB-I (Fiji): Musterung wie Klon B (Ineich, 1988) mit deutlichen lateralen Zeichen am Nacken und unregelmäßigen Flecken (Moritz et al., 1993). Diploide Gruppe Klone 2nB-II (Tahiti): Musterung ähnlich, wie Klon D (Ineich, 1988) allerdings nicht mit der einfachen Reihe weniger Flecken im vorderen Bereich entlang des Rückens sondern eher im hinteren Bereich (Moritz et al., 1993). Triploide Gruppe Klone 3nA (Samoa): Musterung wie Klon A (Ineich, 1988; Moritz et al., 1993). Triploide Gruppe Klone 3nB: Unterschiedlich, manchmal ähnlich Klon B (Ineich, 1988; Moritz et al., 1993). |
| Hybriden: | >> Seitenanfang |
|
Häufig kommt es bei Kreuzungen zwischen Klonen und bisexuellen Tieren zu nicht fortpflanzungsfähigen Hybriden, die weibliche oder männliche Merkmale aufweisen können.
Untersuchungen von Ivan Ineich und Hidetoshi Ota (1992) auf dem Takapoto Atoll ergaben,
dass Hybriden oft deutlich grösser waren als die bisexuellen Lepidodactylus sp. (damals noch ebenfalls als Lepidodactylus lugubris beschrieben) oder die Lepidodactylus lugubris Klone.
Die von Boissinot et al. (1997) auf dem Takapoto Atoll untersuchten sterilen Hybriden wurden ausnahmslos als das Resultat einer Kreuzung zwischen dem diploiden Klon A und Männchen der bisexuellen Lepidodactylus sp. vorort identifiziert. Es wurden keine Tiere gefunden die aus Hybridisierung mit Weibchen von Klon B, C oder D entstanden sein konnten. Boissinot et al. (1997) waren sich nicht sicher, ob dies mit der geringeren Verbreitung dieser Klone oder eventuellen genetischen oder chomosomenbedingten Inkompatibilitäten zu erklären ist oder sonstige Gründe hatte. |
| Klone & Männchen: | >> Seitenanfang |
|
Ohne Fremdeinwirkung schlüpfen aus Eiern parthenogenetischer Geckos für gewöhnlich wieder
parthenogenetische Weibchen.
So stellte sich bei den meisten Berichten über angebliche Männchen der Art später auch heraus, dass es sich um Männchen nahe verwandter Lepidodactylus-Arten oder Hybriden handelte. Dennoch gibt es auch Ausnahmen. Rösler (1992) berichtet von zwei gut entwickelten männlichen Tieren, die bei Terrarienhaltung aus Eiern von Lepidodactylus lugubris schlüpften. Leider gibt er nicht an, ob die Männchen fortpflanzungsfähig waren. Da die Herkunft der Muttertiere nicht geklärt werden konnte, läßt er offen, ob eines der beiden Weibchen bisexuell war, vermutet aber, dass beide parthenogenetisch waren und aus ungeklärten Gründen Männchen schlüpften. Orlando Cuellar und Arnold G. Kluge (1972) berichten, dass die meisten bekannt gewordenen Lepidodactylus lugubris-"Männchen" anomal geformte Hoden mit unüblicher Hautstruktur besitzen, was darauf hinweist, dass die Spermatogenese nicht auf normale Weise erfolgte. Und es gibt noch weitere Punkte, die dafür sprechen, dass diese "Männchen" nicht aus Hybridisierung hervorgegangen sind; insbesondere die Tatsache, dass bei Hybridisierung Phenotypen entstehen, die von der Elternspezies abweichen. Die von Cuellar und Kluge (1972) untersuchten männlichen Lepidodactylus lugubris zeigten aber keinen offensichtlichen Unterschied zum typischen Phenotyp der unisexuellen Weibchen. 1998 beschrieben Yamashiro und Ota ein steriles "Männchen" von der Insel Ishigakijima (Yaeyama Group, Ryukyu Archipel), das genauso aussah wie die dortigen Klon C-Weibchen und aufgrund vorort fehlender verwandter Arten ebenfalls nicht aus Hybridisierung hervorgegangen sein konnte. Sie vermuteten, dass sich das Tier durch hormonelle Veränderungen aus einem normalen Klon C entwickelte. Unter den 673 von Cuellar und Kluge (1972) untersuchten Lepidodactylus lugubris auf Oahu, Hawaii, waren nur 4 Individuen, die männliche Merkmale aufwiesen. Radtkey et al. (1995) nennen sogar nur 5 "Männchen" auf über 8000 Geckos bezogen auf die Hawaiianischen Inseln und Marianas Inseln. Eines meiner eigenen Jungtiere hat sich ebenfalls zu einem "Männchen" entwickelt. Es schlüpfte aus einem Ei, das bei zeitweise um die 30° C im Sommer inkubiert wurde. Ob die hohe Inkubations-Temperatur einen Einfluß auf die Entwicklung hatte oder sonstige Faktoren mitspielen kann ich nicht sagen. |

|
|
Susan G. Brown und Susan Murphy-Walker (1996) dokumentierten das interaktive Verhalten zwischen einem sterilen aber gut entwickelten "Männchen" und jeweils einem von 6 Weibchen in verschiedenen Reproduktions-Stadien: Das Männchen näherte sich Weibchen mit keiner oder geringer Eientwicklung häufiger als den Weibchen mit deutlicher Eientwicklung und wurde beobachtet, wie es Weibchen im Nacken biss und auf sie kletterte, aber es wurde keine Paarung beobachtet. Eventuell fehlte den Weibchen oder dem Männchen das entsprechende Antwort-Verhalten, das zu einer normalen Werbung und Paarung hätte führen können.. Studien von Brown et al. (1993) zeigten, dass sich bei reinen Weibchengruppen eine Rangordnung bildet und sich das interaktive Verhalten reduziert. Die Interaktionen zwischen Männchen und Weibchen blieben jedoch auf hohem Level, wobei die Weibchen aggressiveres Verhalten zeigten als das Männchen. Wie bei anderen Geckospezies reagierte keines der Weibchen auf den Ruf des Männchens, der warscheinlich andere Männchen fernhalten soll (Marcellini, 1977b). |

|
|
Mein Geckomännchen wird von den Weibchen meistens ignoriert. Häufig nähert es sich den Weibchen
mit dem von Brown et al. (1993) beschriebenen "multiple chirp call", der aus mehreren dicht aufeinander
folgenden Lauten, die den Einzellauten der Weibchen ähneln, besteht.
Ein paar mal konnte ich auch beobachten, wie es ein Weibchen am Boden über- und unterkroch und wiederholt im Nacken packte und dann wieder wie unentschlossen los ließ. Das Weibchen blieb dabei unbeeindruckt sitzen, bis es nach einer Weile das aufgeregte Männchen einfach sitzen ließ. |
| >> Seitenanfang |